Eluiga, eriti selle pikkus, on nii teadlastele kui laiemale avalikkusele laialt huvipakkuv teema. Eesti meedias on ilmunud mitmeid eluea seoselisi Tuul Sepa ja Peeter Hõraku populaarteaduslikke artikleid, näiteks siin ja siin. Samas on seda raske uurida, sest eluiga mõjutavad paljud keskkonna- ja pärilikud tunnused, ning neid mõjusid üksteisest eristada ei ole niisama lihtne. Enamasti on erinevate mõjurite eristamisel lahenduseks katsed, kus üht või paari (keskkonna)tegurit manipuleeritakse (näiteks saab kehva toiduga vähendada ka geneetiliselt kiirekasvuliste organismide kasvukiirust ja nii eraldada geneetiline ja keskkonnaseoseline kasvukomponent üksteisest) ja seejärel analüüsitakse seda, mis huvipakkuvate geneetiliste, füsioloogiliste või käitumuslike tunnustega juhtub. Eluea puhul on sellist katset aga keeruline planeerida – ei ole lihtne leida keskkonnategurit, mille muutmine eluiga kindlas suunas mõjutaks. Näiteks kättesaadava toidu hulga vähendamine looteeas või kasvuperioodil võib viia nii eluea pikenemise kui lühenemiseni.
Aasta-ajalised polüfenismid
Ometi on olemas uuritavaid süsteeme, kus üht keskkonnategurit muutes saab esile kutsuda kindlasuunalise ja mastaapse muutuse elueas. Üheks selliseks on aastaajalised polüfenismid: nähtus, kus liikidel, kel esineb aasta jooksul mitu põlvkonda, erinevad põlvkonnad ühe või mitme tunnuse (näiteks muster, värv, kehasuurus, jätkete pikkus jne) poolest. Erinevalt polümorfismist ei ole polüfenism aga geneetiliselt määratud – sama isend võib kasvada ühele või teisele põlvkonnale sarnaseks keskkonnas, kus tavapäraselt vastava põlvkonna isendid elavad. Näiteks mitmete keriloomade ja planktiliste vähkide kehakuju ja jätkete erinevused suviste ja talviste põlvkondade vahel kujunevad eeskätt vastusena veetemperatuuri muutustele, mitte geenidele.
Parasvöötmes elavatel putukatel, kel esineb aastas kaks või enam põlvkonda, tuleb ühel põlvkonnal talv üle elada (nn talvituv põlvkond), samas kui teisel (otsearenev(ad) põlvkon(na)d) seda piirangut ei ole. Kuna putukad (ja muud selgrootud) on kõigusoojased loomad, ei suuda nad olla madalate temperatuuridega aktiivsed, pealegi poleks enamusel neist ka talvisel ajal midagi süüa. Seetõttu on valdav enamus putukatest meie laiuskraadil sügisest kevadeni (s-o rohkem kui poole aastast) inaktiivses olekus ja selle paratamatu järelmina on mitme põlvkonnaga liikidel talvituva põlvkonna eluiga oluliselt pikem kui otseareneval põlvkonnal. Erinevalt keriloomadest ja vesikirbulistest vähkidest kasutavad parasvöötme putukad aga talvitumisotsuse tegemiseks ja põlvkonna eritunnuste väljakujundamiseks valge ja pimeda aja suhet ehk päevapikkust. Viimasel, erinevalt näiteks temperatuurist, elukate arengule enamasti otsest füsioloogilist mõju pole (juhul kui ei ole tegu ekstreemsusega, näiteks alaline pimedus võrrelduna alalise valgusega) ja seetõttu saab mitme põlvkonnaga liblikaid laboris “sundida” arenema talvituva või otseareneva põlvkonna isenditeks muidu identsetes, aga päevapikkuselt erinevates keskkondades. Talvituvale põlvkonnale omased tunnused tekivad siis, kui isendid kasvavad lühikese valguspäeva tingimustes (12 tundi või vähem valgust ööpäevas), otsearenevale põlvkonnale paneb aluse pikk päev. Jõudes tagasi eluea uuringute juurde, on kahe põlvkonnaga liblikaid hea uurida, sest saame mis tahes isendeid füsioloogilise mõjuta päevapikkuse abil panna arenema lühiealiseks otsearenevaks või pikaealiseks talvituvaks vormiks.
Pikema elutsükliga nõgeseliblikas on tugevama tervisega
Koostöös Tartu Ülikooli zooloogia osakonna vilistlase ja nüüdse Greifswaldi Ülikooli (Austria) dotsendi Dalial Freitakiga uurisime nõgeseliblika talvituva ja otseareneva põlvkonna viimase kasvujärgu röövikute immuunvastust nii laboris kasvatatud kui loodusest korjatud isenditel (uuring on siin). Eesmärgiks oli selgitada, kas tunduvalt pikema elueaga talvituva põlvkonna isendid investeerivad lühiealistest otseareneva põlvkonna isenditest rohkem kulukasse immuunsüsteemi, et jääda ellu üsna pika ja muutuva patogeenide koosseisuga elutsükli jooksul. Lühikese elueaga otsearenev põlvkond võiks aga loota haigustekitajate vältimisele ja investeerida immuunsüsteemi arvelt muudesse elukäigutunnustesse, eeskätt kehasuurusesse ja viljakusse, mis on putukatel reeglina omavahel positiivselt seotud. Sellest, kas püstitatud hüpoteesid leidsid kinnitust või mitte, õige pea, aga enne seda põikame uurimuse tausta avamiseks korraks nõgeseliblika bioloogia juurde.
Nõgeseliblika elutsükkel
Nõgeseliblikas on Euraasia parasvöötmes (sh Eestis) laialt levinud ja kliimamuutuste tingimustes kiirelt levilat suurendav päevaliblikas. Suuremas osas levilast on nõgeseliblikal kaks põlvkonda aastas ja ta on üks tuntumaid aastaajalise polüfenismi näiteid – kaks põlvkonda on nii välimuselt kui muudelt tunnustelt väga erinevad (vt joonis 1). Talvituva põlvkonna valmikud (lendavad kevadel) on oranži põhitooniga, väiksemad, pikema nukustaadiumiga (nukk talvitub), vähemarvukamad ja kehvemad levijad. Otseareneva põlvkonna valmikud (lendavad suvel) on aga musta põhitooniga, suuremad, lühema nukustaadiumiga, arvukamad ja paremad levijad. Mõlema põlvkonna liblikatele on iseloomulik munade munemine üksteise otsa, kõrvuti paiknevatesse “tornidesse” (nõgeseõite imiteerimine?, joonis 1) ning viis kasvujärku läbivate ja kõrvenõgesel toituvate, musta või pruuni värvi röövikute koondumine suurtesse salkadesse (joonis 2). Samuti nõgesel toituvatest, musta põhivärviga ning salkadesse koonduvatest päevapaabusilma röövikutest on nõgeseliblika vastseid lihtne eristada (vt joonis 4). Salkadesse koondumise ja suure arvukuse tõttu on nõgeseliblika vastseid loodusest hõlbus leida, kui teada nende peamist põgenemisstrateegiat (taimelt maha kukutamine) ning selle vastu meetmed tarvitusele võtta (vt joonis 4). Eluea uurimise seisukohalt on huvipakkuv just suur vahe eluea pikkuses – eelkõige talvituva nukustaadiumi tõttu kestab Eestis nõgeseliblika talvituva põlvkonna elutsükkel rohkem kui neli korda kauem kui otseareneval põlvkonnal (joonis 1)!





Uurimuse tutvustus, jätk
Kasvatasime osasid nõgeseliblika röövikuid laboris munadest valmikuni (laboriuuring) ning kogusime viimaste kasvujärkude röövikuid ka Tartu ümbrusest (väliuuring). Samaaegset labori- ja väliuuringut ühele uurimisküsimusele vastamiseks kohtab praktilistel põhjustel harva (harva saab seda mugavalt teha nii laboris kui ka metsas); samas on see vajalik, sest laboriuuringute tulemused ei pruugi looduses kehtida (üks näide selle kohta siin) ning loodusest saadud andmete puhul ei ole võimalik erinevate tegurite mõju üksteisest eristada.
Mõlemas uuringus võtsime viimase kasvujärgu röövikutelt hemolümfi (putukate veri) ning mõõtsime sealt kaht immuunparameetrit – lüütilist aktiivsust ja fenooloksüdaasi aktiivsust, mis mõlemad kajastavad lülijalgsete immuunsüsteemi valmisolekut haigustekitajatega võidelda – mida kõrgemad väärtused, seda kõrgem on putuka immunokompetentsus. Lisaks mõõtsime mitmesuguseid muid elukäiguparameetreid nagu kehakaal ja elumus.
Tulemused näitasid selgelt, et tõepoolest on talvituva põlvkonna immunokompetentsus kõrgem kui otseareneval põlvkonnal, seda nii labori- kui välitingimustes (joonis 6) ja lisaks leidis kinnitust tõik, et vastupidiselt tavalisele põlvkondadevahelisele kehasuuruste erinevustele liblikalistel (talvituvad suuremad kui otsearenevad[) on nõgeseliblika talvituva põlvkonna nukud ja valmikud väiksemad (ka laboris eri päevapikkuse juures, kuid muidu samadel tingimustel kasvatades!) kui otseareneva põlvkonna liblikad.
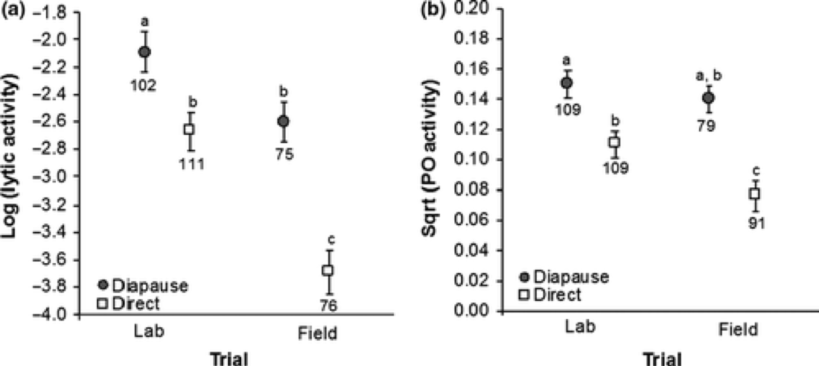
Nagu enamasti ikka, jääb uuringutulemuste tõlgendamiseks mitmeid võimalusi ning nende vähendamiseks oleks vaja teostada uusi uuringuid. Näiteks on võimalik, et hilissuvel, kui talvituv põlvkond röövikupõlve veedab, on haigustekitajaid rohkem kui varasuvel. Ainuüksi sel põhjusel peavad talvituva põlvkonna indiviidid rohkem immuunsüsteemi investeerima. Samas on siiski väga tõenäoline, et talvituva põlvkonna otsearenevast pikem eluiga ja tugevam immuunsüsteem on omavahel põhjuslikult seotud. Seda toetab tulemus, et ka laboris samal temperatuuril, toidul ja patogeenide koosseisu ning arvukuse tingimustes ilmnes samalaadne vahe talvituvate ja otsearenevate isendite immuunvastuses. Talvituva põlvkonna väiksemad mõõtmed võivad aga olla hind, mida rohkem immuunsüsteemi investeerivad isendid maksma peavad – kuluka immuunsüsteemi ülalpidamine ja suurte mõõtmete saavutamine ei pruugi korraga võimalik olla.
Mõistmine, et aastaajalise polüfenismiga liigid kujutavad elueauurijatele kullakaevandust, ning sellesisulised täpsemad uuringud, seisavad aga kindlasti veel ees.
Väga hea! Ainult see hirmus sõna immunokompetentsus…
Aitäh! Nõus, et too monstrum ei kõla hästi ja hea meelega asendaks selle kodumaisema sõnaga, kas sa, Peeter, oskad miskit pakkuda?